 https://orcid.org/0000-0002-6084-7313
https://orcid.org/0000-0002-6084-7313A Descoberta da Estrutura do DNA: de Mendel a Watson e Crick
Originalmente publicado em Química Nova na Escola, n.17, 2003
Apoio: Sociedade Brasileira de Química
Edição: Leila Cardoso TeruyaCoordenação: Guilherme Andrade Marson
Em abril de 2003, o mundo celebrou o cinqüentenário da descoberta da estrutura em dupla hélice do ácido
desoxirribonucléico, mais conhecido como DNA - sua sigla do Inglês
(Watson, 1968; Crick, 1990). A importância dessa descoberta, talvez a
mais importante da história da Biologia moderna, reside no fato do DNA
de cada célula conter toda a informação genética para as características
e funções da mesma. Essa conquista seminal da Ciência está sendo
celebrada de diversas formas por ter atualmente um alcance profundo em
nosso cotidiano. Com o rápido desenvolvimento científico e tecnológico
na área de Biotecnologia, em grande parte iniciado por essa descoberta,
diversos aspectos da sociedade moderna estão sendo afetados.
Esse feito envolveu três instituições de pesquisa. O
Laboratório Cavendish na Universidade de Cambridge, a Unidade de
Biofísica do King’s College, ambos na Inglaterra, e nos Estados Unidos
da América, o Instituto de Tecnologia da Califórnia (mais conhecido pela
sigla Caltech), em Pasadena, onde houve a contribuição do trabalho de
Linus Pauling (1901-1994).
O conceito de que a informação genética das células e
dos organismos vivos que determina as características de cada ser vivo
está contida nas moléculas de DNA é bem aceito na atualidade, mas nem
sempre foi assim. Um grande número de pesquisadores contribuiu para que a
função do DNA fosse finalmente identificada.
|
Primeiros passos
Os
primeiros conceitos de genética, publicados em 1865, foram
desenvolvidos por um monge austríaco, Gregor Mendel (1822-1884), que
trabalhando sozinho deduziu, a partir de experimentos muito bem
elaborados com plantas de ervilha, que algumas características são
herdadas em “unidades” (Figura 1A). Esse trabalho não foi notado pela
comunidade científica até ser descoberto por Hugo De Vries (1848-1935) e
seus colaboradores, em 1900, os quais estabeleceram as leis da
hereditariedade (Figura 1B). Muito tempo se passou e diversas
descobertas foram realizadas descrevendo a composição química das
células.
|

Figura
1: (A) Gregor Mendel e seu jardim no monastério onde realizou os
experimentos de cruzamento com plantas de ervilhas, os quais levaram-no a
desenvolver suas teorias da hereditariedade. (B) Hugo De Vries; em
1900, ele e seus colaboradores redescobriram os trabalhos de Mendel e
formularam as leis da hereditariedade.
|
Poucos
anos após os trabalhos de Mendel, em 1868, um jovem pesquisador,
Friedrich Miescher (1844-1895), purificou uma nova substância no
material nuclear de células, chamada de nucleína (Figura 2). Essa
descoberta não causou grande interesse na época, pois a maioria se
interessava principalmente pelas proteínas que estavam contidas no
núcleo das células.
|

Figura 2: Friedrich Miescher e seu laboratório, onde realizou as purificações e descobriu a nucleína.
|
A
primeira sugestão apontando o DNA como repositório do material genético
de uma célula veio do trabalho de Walther Flemming (1843-1905) que, em
1882, descobriu o processo de mitose e o comportamento dos cromossomos
durante a divisão celular. Essa descoberta, porém, não tornou o DNA o
principal candidato ao posto de carreador da informação genética. Uma
série de descobertas descrevem o DNA como um polímero que contém apenas
quatro formas químicas diferentes: os nucleotídeos adenina (A), guanina
(G), citosina (C) e timina (T) (Figura 3). Em particular, o trabalho de
Phoebus Aaron Theodor Levene (1869- 1940), que fez contribuições
importantes nesse aspecto e, em 1909, propôs a “teoria do
tetranucleotídeo” (Figura 4). Por essa teoria, o DNA seria composto por
repetições monótonas desses quatro nucleotídeos. Dessa forma, uma
molécula com essa característica, certamente não carregaria informação
química suficiente para codificar todas as características necessárias à
manutenção de uma célula e à hereditariedade. Pensava-se que essa
complexidade esperada do material genético seria satisfeita apenas por
proteínas, que são polímeros de 20 aminoácidos.
|
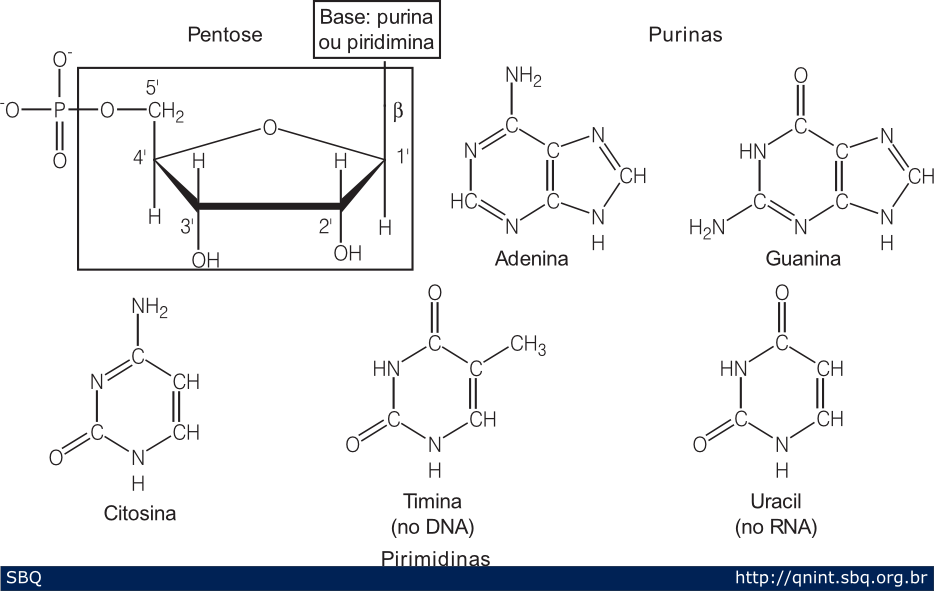
Figura
3: As unidades do DNA são formadas por um nucleotídeo que é composto
por um açúcar (pentose) e uma base nitrogenada. Essa base nitrogenada
pode ser de dois tipos, uma purina ou uma pirimidina. Além do DNA, as
células também possuem moléculas de RNA (sigla do Inglês para ácido
ribonucléico). Este é diferente do DNA por sua molécula ter um outro OH
na posição 2’ da pentose (o DNA possui um H) e, ao invés da base timina
(T), ter a base uracil (U).
|
Figura
4: A teoria do tetranucleotídeo foi introduzida por Phoebus A.T.
Levene, juntamente com inúmeras importantes contribuições de seu
trabalho na caracterização química do DNA.
|
Apesar
disso, a pesquisa com o DNA prosseguiu e, em 1909, um geneticista
dinamarquês, Wilhelm L. Johannsen (1857-1927), cunhou o nome “gene” para
a unidade descrita por Mendel e, em 1911, Thomas Hunt Morgan
(1866-1945), trabalhando com a mosca da fruta, Drosofila melanogaster,
mostrou pela primeira vez que os genes estão arranjados de forma linear
nos cromossomos (Figura 5).
|

Figura
5: (A) Wilhelm Johannsen fotografado durante uma apresentação a seus
alunos; foi o responsável por cunhar o nome “gene” para a unidade de
transferência da hereditariedade. (B) Thomas H. Morgan; trabalhando com
moscas de fruta (Drosophila melanogaster), identificou que os genes estão arranjados em seqüência nos cromossomos.
|
Evidências sobre o papel do DNA
As
evidências experimentais de que o DNA é o material genético surgiram de
uma série de experimentos muito elegantes realizados a partir de 1928.
Nesse ano, Frederick Griffith, usando extratos químicos, converteu
inócuas bactérias de pneumonia na sua forma patogênica. A natureza desse
fator de hereditariedade não foi sugerido por ele na ocasião.
Em um interessante livro de 1943, intitulado What is
life?, Erwin Schrödinger (1887-1961) apresentou o conceito dos genes
serem a componente chave das células vivas ( Schrödinger, 1997).
Entretanto segundo os paradigmas da época, ele supôs que os genes seriam
uma classe especial de proteínas. Mas logo no ano seguinte, em plena 2ª
Guerra Mundial, Oswald T. Avery (1877-1955) e seus colegas, Colin
MacLeod (1909-1972) e Maclyn McCarty (1911-) - Figura 6, baseados nas
observações de Griffith, demonstraram ser o DNA a molécula responsável
pelo princípio transformante. O experimento definitivo surgiu somente em
1952, com os elegantes experimentos de Alfred Hershey (1908-1997) e
Martha Chase (1930-) - Figura 7. Dessa forma, ficou consolidada a noção
de que o material genético é constituído de moléculas de DNA e não de
proteínas, como alguns pesquisadores ainda acreditavam até então. Com
isso se tornou de grande importância a descoberta da estrutura dessa
intrigante molécula, pois já se especulava que uma descoberta dessa
natureza revelaria como a informação é armazenada no DNA e como ela
seria transmitida de um indivíduo para seu descendente.
|

Figura
6: Oswald T. Avery (esq.) e seus colaboradores Colin MacLeod e Maclyn
McCarty demonstraram o princípio transformante proposto por Frederick
Griffith em 1928.
|
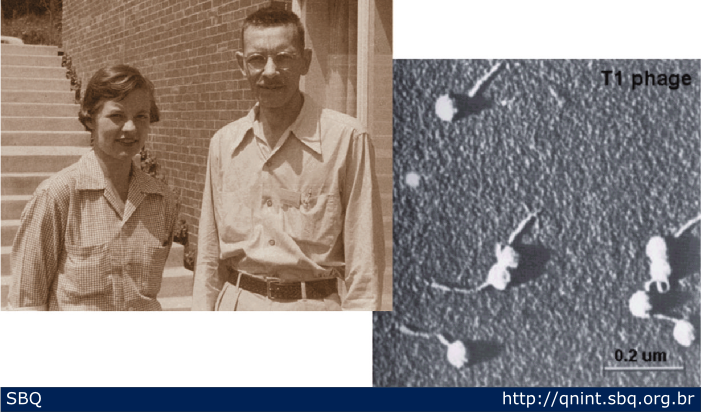
Figura
7: Martha Chase e Alfred Hershey; em seus experimentos com
bacteriófagos, vírus que infectam bactérias, demonstraram
definitivamente que o DNA é o material responsável pela transmissão dos
caracteres hereditários.
|
Uma
descoberta fundamental para tal foi realizada por Erwin Chargaff
(1905-2002), em 1950 (Figura 8). Chargaff investigou a composição do DNA
de diversos organismos e concluiu que a composição de bases do DNA
varia entre as espécies, mas que amostras de DNA isoladas de diferentes
tecidos da mesma espécie têm a mesma composição de bases (A, C, T e G).
Ele também pôde observar que a composição de bases do DNA numa dada
espécie não muda com a idade do organismo, estado nutricional ou
mudanças ambientais. Sua observação mais importante, porém, talvez tenha
sido de que em todos os DNA celulares, não importa qual a espécie, o
número de bases adenina é igual ao de timina (A = T) e o de guanina é
igual ao de citosina (G = C). Essa passou a ser conhecida como “Regra de
Chargaff” (Figura 8).
|

Figura
8: Erwin Chargaff; após inúmeros estudos da composição do DNA em
diversos tecidos e varias espécies, concluiu que a ocorrência das quatro
bases no DNA obedece às relações A = T e C = G. Esta regra é conhecida
como “Regra de Chargaff”.
|
Passos para a descoberta da estrutura do DNA
Entre
1944 e 1945, Maurice Wilkins (1916-) - Figura 9 - trabalhava em
processos de separação de isótopos de urânio com espectrógrafos de massa
no Projeto Manhattan, em Berkeley (EUA). Nesse período, leu o livro de
Schrödinger e se interessou pelo tema. Depois de ler os artigos
publicados pelo grupo de Avery, Wilkins supôs que o DNA era o
transmissor da hereditariedade e, em 1947, foi para Londres trabalhar
com John T. Randall (1905- 1984) - Figura 9 - sobre vários temas ligados
ao DNA, na récem-criada Unidade de Biofísica do King’s College. Iniciou
os primeiros experimentos de difração de raios X com fibras de DNA em
1950, depois de receber uma pequena quantidade de DNA muito puro de
Rudolph Signer, um bioquímico suíço.
|

Figura
9: Rosalind E. Franklin trabalhou com Maurice Wilkins no estudo do DNA.
Wilkins e John Randall trabalharam juntos na Unidade de Biofísica do
King’s College em Londres.
Os
experimentos de difração de raios X realizados no laboratório de
Wilkins foram conduzidos por Rosalind Elsie Franklin (1920-1958) -
Figura 9, que se juntou à Unidade de Biofísica do King’s College em
novembro de 1951 para trabalhar com DNA (sobre Franklin, vide Farias,
2001 ou Maddox, 2002). Ela produziu fibras muito finas de DNA e as
irradiou com um feixe ultrafino de raios X. Em pouco tempo ela descobriu
que o DNA se apresentava em duas formas diferentes, as quais denominou
de A e B (Figura 10). A forma A, facilmente fotografada, correspondia a
fibras de DNA desidratadas, enquanto a forma B correspondia a fibras
molhadas de DNA. Apesar de mais difícil de registrar com os raios X, a
forma B mostrava um padrão compatível com uma hélice (Figura 10). Uma
vez que a água poderia ser atraída pelos grupos fosfato do DNA, e este
poderia ser facilmente hidratado e desidratado, ela sugeriu que os
fosfatos do DNA se localizavam no exterior da hélice e as bases
nitrogenadas (A, C, G e T) estariam assim voltadas para o interior.
|
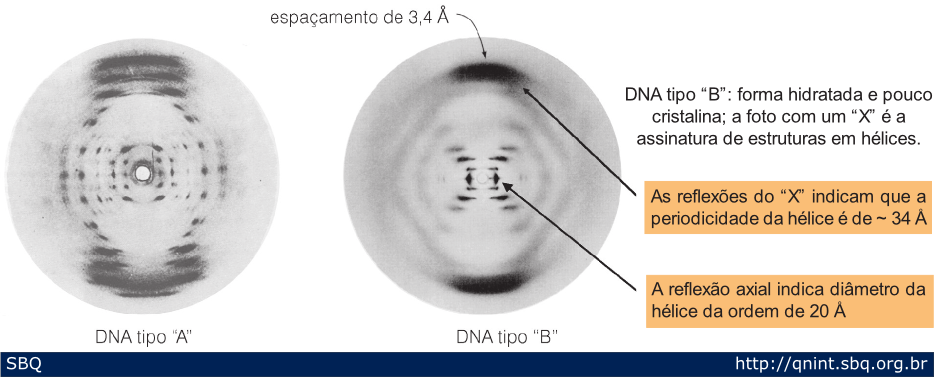
Figura
10: Duas fotos de difração de raios-X obtidas por Rosalind ranklin e
Maurice Wilkins do DNA desidratado (A) e hidratado (B). O exame da foto
de difração do DNA B permitiu a Watson e Crick concluírem diversas
características da hélice.
|
Ao
mesmo tempo, no Caltech, Pauling (Figura 11) também estava trabalhando
para resolver a estrutura do DNA, com o uso de poucos dados
experimentais: apenas algumas fotos de difração de fios de cabelo e seu
aguçado bom-senso químico. Essas duas ferramentas, associadas ao uso de
modelos moleculares, levaram Pauling a deduzir a estrutura em hélice
alfa para as proteínas.
|
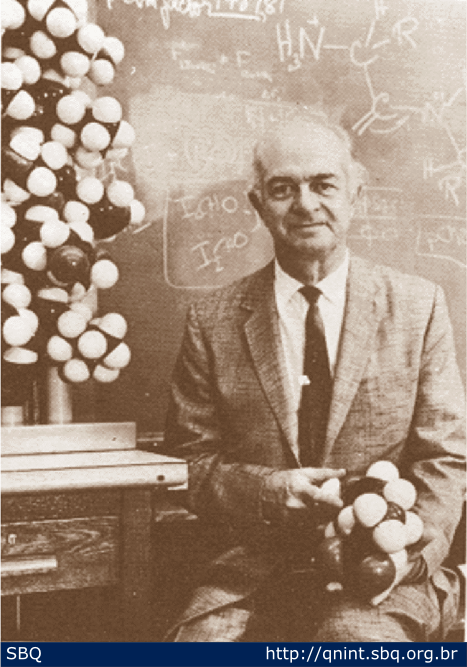
Figura
11: Linus Pauling, fotografado com modelos da estrutura alfa hélice de
proteínas, também se dedicou a resolver a estrutura do DNA.
|
A descoberta da estrutura do DNA
Nesse
mesmo ano, Francis Harry Crick (1916-) - Figura 12 - estava trabalhando
em sua tese de doutorado no Laboratório Cavendish, estudando a
estrutura cristalina da hemoglobina, sob orientação de Max Perutz (1914-
2002). Crick se formou em Física e durante a 2ª Guerra trabalhou para o
Almirantado Britânico, indo estudar Biologia em 1947. Em 1949 se juntou
à unidade chefiada por Max Perutz e sir William Lawrence Bragg
(1890-1971) - Figura 12. Nesse período, Crick conheceu James Dewey
Watson (1928-) - Figura 12 -, que muito influenciou sua carreira.
|

Figura
12: Francis Crick e James Watson foram trabalhar juntos em Cambridge
sob a supervisão de Max Perutz e John Kendrew, respectivamente, no grupo
liderado por sir William Lawrence Bragg.
|
Watson,
formado em Biologia e com um doutorado em Zoologia estudando o efeito
de raios X na multiplicação de bacteriófagos (vírus que infectam
bactérias), realizou seu primeiro ano de pós-doutorado em Copenhagen.
Nessa época, conheceu Wilkins em um simpósio em Nápoles. Ao assistir a
palestra de Wilkins, ficou estimulado pelas imagens de difração de DNA e
decidiu mudar o rumo de sua pesquisa. No outono de 1951, mudouse para o
Laboratório Cavendish, trabalhando sob a supervisão de John C. Kendrew
(1917-1997) – Figura 12. Em pouco tempo Crick e Watson se tornaram
amigos e descobriram seu interesse comum pela estrutura do DNA. Eles
consideraram que seria possível resolver a estrutura do DNA baseados nas
imagens de difração obtidas por Franklin e Wilkins, do King’s College, e
por um cuidadoso exame das configurações estereoquímicas da cadeia de
polinucleotídeos. A abordagem seria semelhante à usada por Pauling,
usando modelos e dedução lógica. A primeira investida de Watson e Crick
para resolver a estrutura foi naquele mesmo outono de 1951. Crick se
encarregou de realizar os cálculos teóricos fundamentais sobre a
difração de hélices. Nessa mesma época Watson foi assistir a um
seminário de Franklin no King’s College sobre seus dados de difração;
porém, por não tomar nota dos dados apresentados por ela, errou a
quantidade de água estimada por Franklin nas fibras de DNA. Baseados
nessa informação incorreta, eles decidiram que a hélice de DNA deveria
ser composta por três cadeias de nucleotídeos. Para poderem obter a
regularidade de hélice observada nos experimentos de difração, decidiram
que os fosfatos estariam posicionados para o interior da estrutura e
não, para o exterior. Para compensar as cargas negativas dos fosfatos,
adicionaram ao modelo íons magnésio no interior da hélice (Figura 13).
|
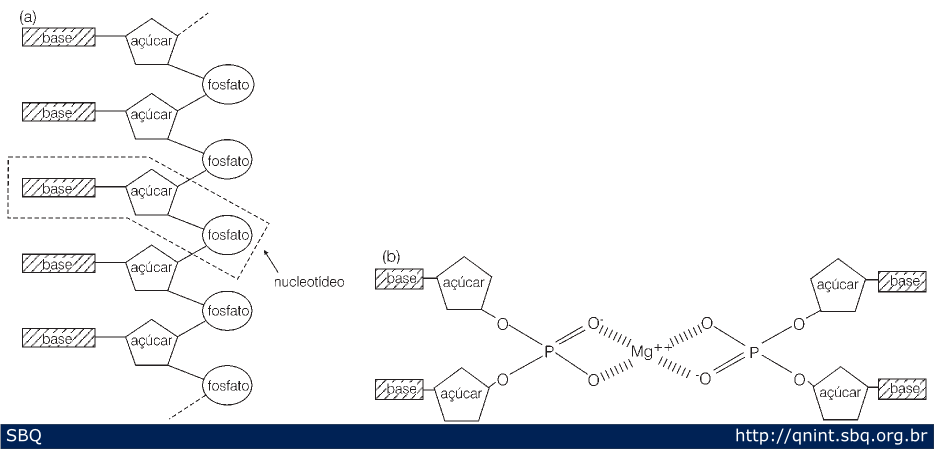
Figura
13: (A) Se sabia que o DNA era um polímero de nucleotídeos, onde um
nucleotídeo e o seguinte são unidos por uma ligação tipo fosfodiester.
Esse fosfato confere a característica ácida ao DNA. (B) Primeira
proposta de Watson e Crick para a estrutura do DNA: uma tripla hélice
com os fosfatos no interior e as bases para fora (no modelo, para
compensar as cargas negativas dos fosfatos, eles introduziram íons de
magnésio).
|
No
ano seguinte, o filho de Linus Pauling, Peter, foi para Cambridge fazer
doutorado com Kendrew. Por intermédio dele, Watson e Crick ficaram
sabendo de um artigo de Pauling descrevendo a estrutura do DNA como uma
tripla hélice, à semelhança da estrutura em que estavam trabalhando.
Rapidamente perceberam o erro, pois os fosfatos para o interior da
estrutura os tornariam protonados e assim o DNA perderia o seu caráter
ácido.
Esse erro de Pauling estimulou-os a retomarem com
vigor essa linha de pesquisa, pois sabiam que não demoraria muito para
Pauling perceber o seu erro e corrigi-lo. Em uma discussão com Wilkins
no King’s College sobre o artigo de Pauling, Watson viu novamente as
figuras de difração do DNA na forma hidratada (B), obtidas por Franklin,
e ficou sabendo de sua conclusão de que os fosfatos deveriam estar do
lado de fora da hélice.
Depois de seguidas tentativas, em 28 de fevereiro de
1953 Watson fez modelos das bases (A, C, G e T) em pedaços de cartão
(Figura 14), na tentativa de identificar possíveis modos de interação.
Percebeu então que os pares A - T e C - G formavam ligações de
hidrogênio, resultando em pares de dimensões quase idênticas, o que
permitiria que a hélice se mantivesse com o mesmo diâmetro, independente
do pareamento de bases no interior. Esse arranjo satisfez à regra
proposta por Chargaff, pela qual A = T e C = G. Depois de trabalharem
sobre esse modelo por mais alguns dias, refinando o mesmo para que fosse
coerente com os dados de difração de raios X, eles chegaram ao modelo
final (Figura 14).
|
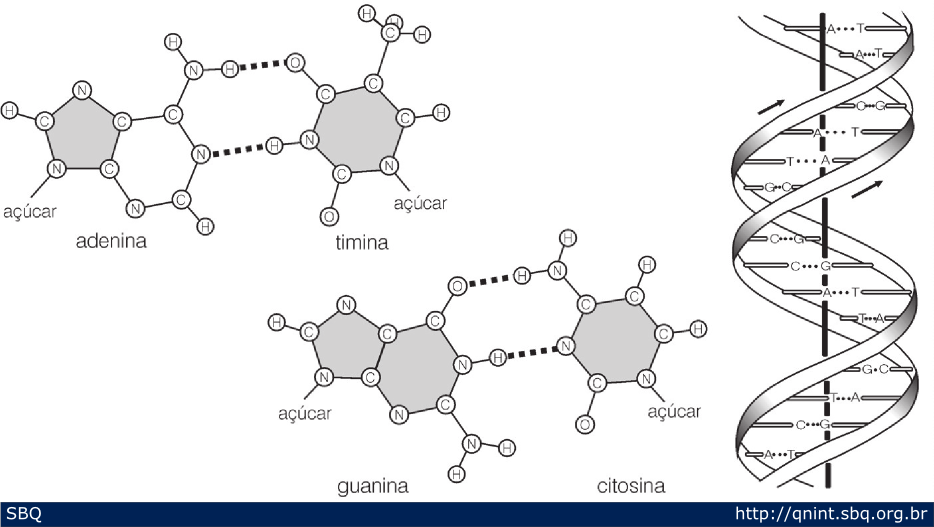
Figura
14: Empregando as bases desenhadas em cartões, Watson percebeu como
deveriam se formar pontes de hidrogênio entre elas e a importância da
relação de Chargaff. Nesta figura está ilustrada a primeira hipótese na
qual o número de pontes de hidrogênio entre C e G é de apenas duas.
Posteriormente verificaram a possibilidade de se formarem três pontes
entre C e G. Finalmente, após alguns refinamentos no modelo, chegaram à
proposta final da estrutura do DNA.
|
Pouco
depois, em março, Wilkins e Franklin visitaram o laboratório onde
trabalhavam Watson e Crick para ver o modelo (Figura 15). Nessa ocasião,
Franklin mostrou seus dados que inquestionavelmente posicionavam os
fosfatos para fora da hélice. Em uma visita subseqüente de Linus
Pauling, eles mostraram o modelo que foi rapidamente aprovado por
Pauling. Em 2 de abril de 1953 submeteram seu modelo da estrutura do DNA
em um artigo para a renomada revista Nature. A esse artigo seguiu-se a
proposta de um esquema de replicação da molécula de DNA (mostrado na
Figura 15, à direita).
|
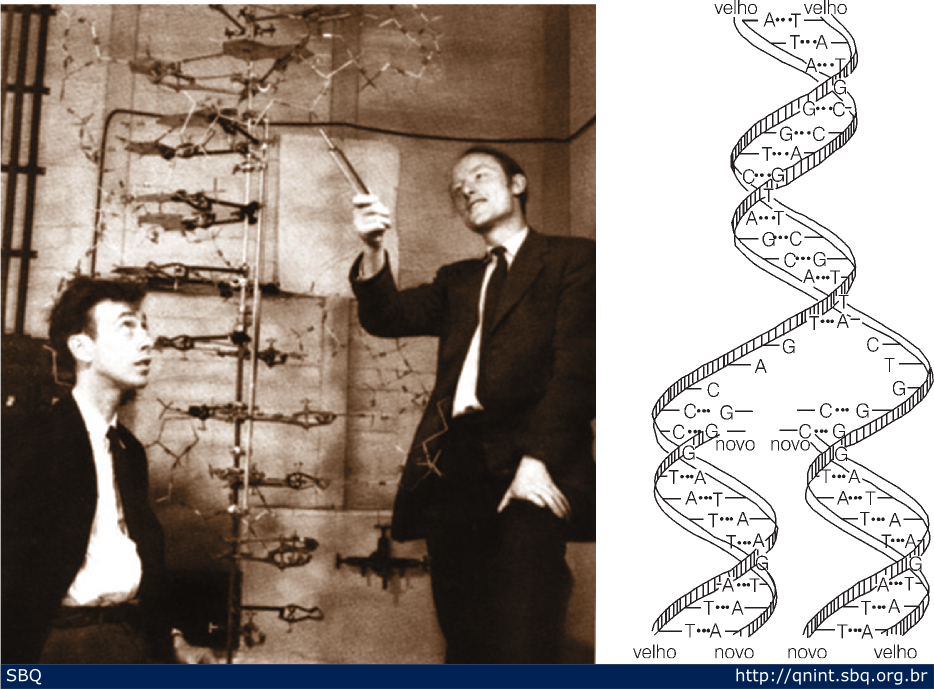
Figura
15: Famosa fotografia de Watson e Crick explicando seu modelo aos
visitantes do laboratório. A informação de Franklin de que os fosfatos
deveriam estar do lado de fora da molécula foi crucial para a montagem
do modelo.
|
No
ano seguinte, Crick obteve seu título de doutor com a tese “Difração de
raios X: polipeptídeos e proteínas”. Mais tarde, e em colaboração com
Sydney Brener, Crick contribuiu com avanços fundamentais no campo da
síntese de proteínas e do código genético.
Em 1962, Crick, Watson e Wilkins compartilharam o
Prêmio Nobel de Medicina ou Fisiologia pela descoberta da estrutura do
DNA. No mesmo ano, Perutz e Kendrew dividiram o Prêmio Nobel de Química
pelos estudos estruturais de proteínas globulares (Figura 16).
|

Figura
16: Foto de Maurice Wilkins, John Steinbeck, John Kendrew, Max Perutz,
Francis Crick e James Watson, quando do recebimento dos Prêmios Nobel de
Medicina ou Fisiologia, de Literatura e de Química, em 1962.
|
O
modelo proposto por Watson e Crick explica perfeitamente os dados de
difração de raios X e permite deduzir a forma de replicação da molécula
de DNA. Representa até hoje um marco na história da Ciência, permitindo a
fundação da área de pesquisa hoje conhecida como Biologia Molecular, na
qual a manipulação do DNA recombinante tem papel fundamental.
|
- Referências
- CRICK, F. What mad pursuit: a personal view of scientific discovery. Nova Iorque, HarperCollins Publishers, 1990.
- FARIAS, R.F. de. As mulheres e o Prêmio Nobel de Química. Química Nova na Escola, n. 14, p. 28-30, 2001.
- MADDOX, B. Rosalind Franklin: the dark lady of DNA. Nova Iorque, HarperCollins Publishers, 2002.
- SCHRODINGER, E. O que é vida? O aspecto físico da célula viva seguido de mente e matéria e fragmentos autobio- gráficos. Trad. J. de P. Assis e V.Y.K. de P. Assis. São Paulo: Fundação Editora da Unesp, 1997.
- WATSON, J.D. The double helix: a personal account of the discovery of the structure of DNA. Nova Iorque, The New American Library, 1968.
- FERREIRA, R. Watson & Crick. A história da descoberta da estrutura do DNA. São Paulo: Odysseus, 2003.
- HAUSMAN, R. História da Biologia Molecular. Ribeirão Preto, Funpec Editora, 2002.
- STRATHERN, P. Crick, Watson e o DNA em 90 minutos. Trad. M.L.X.A. Borges. Rio de Janeiro: Jorge Zahar, 2001.